- Barajar
ActivarDesactivar
- Alphabetizar
ActivarDesactivar
- Frente Primero
ActivarDesactivar
- Ambos lados
ActivarDesactivar
- Leer
ActivarDesactivar
Leyendo...
Cómo estudiar sus tarjetas
Teclas de Derecha/Izquierda: Navegar entre tarjetas.tecla derechatecla izquierda
Teclas Arriba/Abajo: Colvea la carta entre frente y dorso.tecla abajotecla arriba
Tecla H: Muestra pista (3er lado).tecla h
Tecla N: Lea el texto en voz.tecla n
![]()
Boton play
![]()
Boton play
![]()
36 Cartas en este set
- Frente
- Atrás
|
Glucógeno
|
• Es un polisacárido ramificado de glucosa
• Está compuesto de cadenas de unidades glucosilo unidas por enlaces α-1,4 con ramificaciones α-1,6 cada 8-10 unidades |
|
Carbono anomérico
|
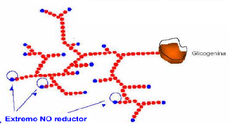
• En esta molécula tan ramificada sólo un residuo de glucosa presenta su C anomérico libre (no está unido a otro residuo de glucosa)
• Este residuo de glucosa, ubicado al principio de de la cadena se encuentra unido a la proteína glucogenina por su C anomérico. • A diferencia de este residuo de glucosa, los ubicados en los extremos de las ramificaciones son no reductores ya que su C anomérico forma parte de un enlace glucosídico |
|
Ventajas de la estructura ramificada del glucógeno
|
Permite su síntesis y degradación rápida, dado que las E que lo metabolizan pueden actuar sobre muchas cadenas simultáneamente (ya que actúan sobre los múltiples extremos no reductores)
|
|
Distribución del glucógeno
|
En alguna medida, el glucógeno está presente en todos los tipos celulares, pero nos centraremos en el músculo esquelético y en el hígado
|
|
Representación del peso del glucógeno
|
• El glucógeno representa el 10% de su peso
• En el músculo representa el 1 o 2% del peso del tejido, pero al ser mayor la masa muscular, los depósitos de glucógeno del músculo son casi el doble que los hepáticos |
|
Degradación del glucógeno
|
• Aporta principalmente glucosa-1-fosfato que se puede convertir en glucosa-6-fosfato
• En el músculo esquelético y en otros tejidos, la glucosa-6-fosfato entra en la vía glucolítica • El glucógeno es una fuente de combustible extremadamente importante para el músculo cuando la demanda de ATP es elevada y cuando se utilizan rápidamente la glucosa-6-fosfato en la glucólisis anaeróbica • En general en estos tejidos la glucogenólisis y la glucólisis se activan simultáneamente |
|
Glucógeno en el músculo (fibras musculares rojas)
|
• El ejercicio activa la movilización del glucógeno muscular para la formación de ATP
• Las fibras musculares rojas reciben un buen flujo sanguíneo, contienen elevados niveles de mioglobina y una gran cantidad de mitocondrias • El glucógeno en estas células se metaboliza a piruvato y dada la presencia de O2 y mitocondrias, el piruvato puede oxidarse a CO2 y H2O |
|
Glucógeno en el músculo (fibras musculares blancas)
|
• Las fibras musculares blancas tienen un flujo sanguíneo menor y menos mitocondrias.
• En estas células, el glucógeno aporta sustrato para la glucólisis, siendo el lactato el producto principal • Las fibras musculares blancas tienen una mayor capacidad de glucogenólisis y glucólisis que las fibras rojas |
|
Glucógeno en el hígado
|
• En el hígado, el glucógeno es la primera y más directa fuente de glucosa para el mantenimiento de la glucemia
• En el hígado, la glucosa-6-fosfato, producto de la degradación del glucógeno, se hidroliza a glucosa por la glucosa-6-fosfatasa, una enzima presente sólo en hígado y riñón • El glucógeno, por lo tanto, es una fuente rápidamente movilizable de glucosa para la sangre, cuando el aporte de glucosa de la dieta disminuye o cuando el ejercicio incrementa su utilización por los músculos |
|
Glucogenólisis y gluconeogénesis
|
• La glucogenólisis y gluconeogénesis hepática aportan glucosa a la sangre y, consecuentemente estas 2 vías se activan simultáneamente
• La gluconeogénesis también produce glucosa-6-fosfato |
|
Niveles de glucógeno hepático
|
• Varían en respuesta a la ingesta de alimentos
• Aumentan inmediatamente después de una comida • Disminuyen lentamente a medida que se moviliza el glucógeno para mantener la glucemia • Esta reserva se utiliza principalmente entre las comidas y aún más durante el ayuno nocturno • En los humanos el glucógeno hepático dura entre 12 y 24 horas de ayuno, dependiendo de la actividad realizada por el individuo |
|
Paso 1 de la síntesis del glucógeno
|
• Comienza con la fosforilación de la glucosa
• Glucosa + ATP -----> Glucosa-6-fosfato + ADP • Hexoquinasa o glucoquinasa • Cofactor: Mg2+ |
|
¿La glucosa-6-fosfato es sustrato de qué vías?
|
• Glucólisis
• Vía de las pentosas • Síntesis de otros azúcares |
|
Paso 2 de la síntesis de glucógeno
|
• Glucosa-6-fosfato <-----> Glucosa-1-Fosfato
• Fosfoglucomutasa • Cofactor: Mg2+ |
|
Paso 3 de la síntesis de glucógeno
|
• Glucosa-1-Fosfato + UTP -----> UDP-Glucosa + PPi
• Glucosa-1-P uridiltransferasa • Cofactor: Mg2+ |
|
Reacción de la pirofosfatasa
|
• La síntesis de UDP-glucosa se desplaza hacia la formación del producto, o sea que se hace energéticamente favorable e irreversible debido a que el pirofosfato producido es hidrolizado subsecuentemente por la pirofosfatasa:
• Pirofosfato (PPi4-) + H2O -----> 2 Fosfatos (Pi2-) |
|
Degradación del glucógeno
|
• Los enlaces glucosídicos si clivan (cortan) por la adición de un fosfato (fosforólisis) para producir glucosa-1-P (o agua para producir glucosa libre), y no se vuelve a sintetizar UDP-Glucosa
|
|
Función de las vías separadas de la síntesis y degradación del glucógeno
|
• La existencia de vías separadas para la formación y degradación de compuestos es un punto clave en el metabolismo.
• Debido a que la síntesis y la degradación utilizan diferentes enzimas es posible activar una vía e inhibir simultáneamente la contraria |
|
Síntesis de glucógeno
|
• En su mayor parte la síntesis de glucógeno consiste en el alargamiento de cadenas de moléculas de glucógeno preexistente (primer o cebo de glucógeno) en la cual el extremo reductor está unida a la proteína glucogenina
|
|
Alargamiento de la cadena de glucógeno
|
• Para alargar las cadenas, la enzima glucógeno sintasa agrega residuos de glucosa a partir de UDP-glucosa a los extremos no reductores de la cadena
• El C anomérico (C1) de cada residuo de glucosa que se incorpora se une por un enlace α-1,4 al hidroxilo (OH) del C4 del último residuo de glucosa de la cadena |
|
Reconversión de UDP
|
•El UDP puede reconvertirse en UTP
• UDP + ATP -----> UTP + ADP • Nucleósido difosfato quinasa |
|
Paso regulatorio de la vía
|
La reacción catalizada por la glucógeno sintasa
|
|
11 unidades de glucosa de longitud
|
• Cuando la cadena alcanza 11 unidades de glucosa de longitud, una enzima denominada amilo-4:6-transferasa (enzima ramificante) corta un fragmento de 6 a 8 unidades del extremo no reductor y lo une a un residuo de glucosa por un enlace α-1,6
• La distancia entre 2 ramificaciones consecutivas es de por lo menos 4 residuos de glucosa |
|
Balance completo de la síntesis de glucógeno
|
(Glucosa)n + Glucosa + 2 ATP --> (Glucosa)n+1 + 2 ADP + 2 Pi
|
|
¿Por qué es necesario que existan depósitos de combustible en forma de glucógeno en vez de almacenar esas calorías en forma de lípidos?
|
1) Los lípidos almacenados no pueden degradarse tan rápidamente como el glucógeno
2) en ausencia de oxígeno no se pueden utilizar los lípidos como fuente de energía 3) los lípidos no pueden convertirse en glucosa y por lo tanto no pueden utilizarse para mantener la glucemia (algunos tipos celulares sólo utilizan glucosa como fuente de energía). |
|
¿Por qué es necesario gastar ATP para sintetizar una molécula enorme y compleja como el glucógeno en vez de almacenar la glucosa como tal?
|
1) Por una parte, la glucosa es osmóticamente activa y se necesita ATP para mantenerla dentro de la célula.
2) Por otra, para alcanzar la cantidad de glucosa que se almacena como glucógeno se debería llegar a una concentración intracelular de glucosa de 400 mM lo que produciría la entrada de agua y finalmente ocasionaría la lisis osmótica de las células. |
|
Síntesis de nuevas moléculas de glucógeno
|
• Este proceso ocurre cuando la glucogenina, proteína a
la cual se une el glucógeno, se glicosila (autoglicosilación) uniéndose una molécula de glucosa al OH- de una tirosina. • La adición de residuos de glucosa continúa hasta que la cadena glucosídica es suficientemente larga como para ser sustrato de la glucógeno sintasa. |
|
¿Cuándo termina la síntesis de glucógeno?
|
El glucógeno mismo inhibe a la glucógeno sintasa y por lo tanto evita su síntesis continua.
|
|
Degradación del glucógeno
|
El glucógeno se degrada por la acción combinada de dos enzimas, la glucógeno fosforilasa y la enzima desramificante.
|
|
Degradación del glucógeno (fosforilasa)
|
• La fosforilasa inicia su actividad en el extremo no
reductor de una cadena y libera secuencialmente residuos de glucosa-1-fosfato. • Es una reacción de fosforólisis donde se incorpora un grupo fosfato al carbono anomérico (C1) del último residuo de glucosa de la cadena. • Sin embargo, la fosforilasa no puede actuar sobre los enlaces glucosídicos cuando la degradación llega a 4 residuos de un punto de ramificación, porque ésta impide estéricamente la ubicación correcta de la molécula en el sitio catalítico de la enzima. |
|
Degradación del glucógeno (enzima desramificante)
|
• La enzima desramificante cataliza la remoción de esos 4 residuos.
• Esta enzima presenta dos actividades catalíticas: - --- Actúa como 4:4 transferasa - y como 1,6 glucosidasa • Como transferasa remueve primero una unidad de 3 residuos de glucosa y los agrega al final de otra cadena mediante un enlace α-1,4. • El residuo de glucosa remanente en la ramificación se hidroliza por la actividad amilo-1,6-glucosidasa liberándose como glucosa. • Por lo tanto, en cada punto de ramificación se liberan una molécula de glucosa y alrededor de 7 a 9 de glucosa-1-fosfato. |
|
Siguiente paso en la degradación del glucógeno
|
• Glucosa-1-P <-----> Glucosa-6-P
• Fosfoglucomutasa • Esta reacción, en las condiciones que se encuentran dentro de las células, está casi en equilibrio, lo que le permite funcionar tanto en la síntesis como en la degradación del glucógeno. |
|
Siguiente paso de la degradación de glucógeno (en hígado)
|
• glucosa-6-fosfato+ H2O -----> glucosa + Pi
• Glucosa-6-fosfatasa |
|
Balance de la degradación de glucógeno
|
(glucosa)n + H2O -----> (Glucosa)n-1 + glucosa
No se utiliza ni se forma ATP en este proceso |
|
Siguiente paso de la degradación de glucógeno (en el músculo)
|
• En el músculo la glucosa-6-fosfato se utiliza en la vía glucolítica que lleva principalmente a la producción de lactato en las fibras musculares blancas y a la oxidación
completa de la glucosa en las rojas. |
|
Degradación lisosomal del glucógeno
|
• La degradación del glucógeno también ocurre, en parte, dentro de los lisosomas cuando las partículas de glucógeno se rodean por membranas que luego se fusionan con las membranas lisosomales.
• Una glucosidasa lisosomal hidroliza el glucógeno a glucosa. |